The purpose of this experiment was to determine whether image inversion disrupted the processing of face stimuli more than the processing of landscape stimuli. The experimental hypothesis was that stimulus inversion will impair the recognition of face stimuli more than landscape stimuli. 12 males and 12 females were randomly allocated into either a “face” or “landscape” group, with each group containing 12 participants.
All participants were Level 2 psychology students aged 18-40 from the University of Aberdeen, Scotland. During the testing phase of the experiment, participants from the face group were shown 12 upright images of male faces for 6 seconds per image and then shown 24 upright images of male faces containing 12 target faces and 12 distracter faces and were asked to write down whether they had previously seen the face. This procedure was repeated for the second phase of the experiment with inverted images of male faces.
The exact same procedure was also carried out with upright and inverted landscape images with participants in the “land” group. The results from this experiment showed that image inversion disrupted the processing of face stimuli more than the processing of landscape stimuli, and that these results were statistically significant. On the basis of these findings, the experimental hypothesis was accepted.
Introduction
Face perception involves the processing of the characteristics and features of a face, which in turn allows one to distinguish between and recognise different individuals. Facial recognition is therefore an important and integral part of everyday life, as without it, one may lose the ability to identify familiar faces such as those of relatives or friends.
Individuals with prosopagnosia display such a disorder, as sufferers find it difficult to distinguish between different human faces. However, the ability of prosopagnosics to identify objects (Duchaine and Nakayama, 2005), suggests that there may be a specific region of the brain devoted to facial perception and processing. Some studies suggest that this region may lie in the right hemisphere of the brain.
McNeil and Warrington (1993) for example, who carried out research on a 51 year old man named W.J who had suffered damage to the right hemisphere as a result of a stroke, found that even though W.J was unable to distinguish between pictures of different human faces, he was able to distinguish between pictures of different sheep faces.
Another individual, this time a 33 year old woman named B.M, a right hemispherectomy patient since age 13, had similar difficulties in recognising familiar faces and was unaware that a person could be identified solely by looking at their face as opposed to looking at what they were wearing (Sergent and Villemure, 1989).
Furthermore, the ability to identify a face appears to involve the use of a different brain region than that used to identify facial expressions. Some prosopagnosics for example, can judge emotional states based on a person’s facial expression. While some sufferers of dementia, who are able to recognise faces, have difficulty recognising facial expressions (Kurucz and Feldmar, 1979).
In terms of a specific brain region that is used for face recognition, fMRI studies suggest that the occipital face area (OFA) in the inferior occipital cortex may be involved. This was shown by the case of a 41 year old prosopagnosic woman named P.S who suffered damage to this area of the brain, yet maintained an intact right fusiform gyrus and fusiform face area (FFA) (Rossion, Caldara, Seghier, Schuller, Lazeyras, and Mayer, 2003).
Further evidence to support the role of the OFA in face recognition comes from patient D.F, who, like patient P.S, had an intact FFA but no OFA and was unable to identify familiar faces (Steeves, Culham, Duchaine, Pratesi, Valyear, Schindler, Humphrey, Milner and Goodale, 2006).
The existence of a dedicated face processing region in the brain appears to be both present and active from birth. Maurer and Barrera (1981) for example, found that at two months of age babies spent more time looking at pictures of faces than they did at scrambled or upside down faces, thus showing that they could discriminate amongst them.
At ages three to six months, babies were able to discriminate between different facial expressions (Barrera and Maurer, 1981) and even between attractive and unattractive faces (Samuels and Ewy, 1985) spending more time looking at the former than they did at the later (Langlois, Ritter, Roggman, and Vaughn, 1991).
If, as evidence suggests, there is a specific region of the brain devoted to face recognition, it is not unreasonable to assume that it is designed to process upright faces as this is the orientation in which faces normally appear in everyday life.
Individuals without any disorders to this region should therefore find it easier to identify upright faces as opposed to inverted faces. Conversely, individuals suffering from prosopagnosia as a result of damage to this face recognition region should find it more difficult to process upright faces than they do inverted faces.
Research carried out by Farah, Wilson, Drain, and Tanaka (1995) does seem to support these assumptions, as it was found that normal test subjects were better able to recognise upright faces than inverted faces. However, a prosopagnosic patient, L.H, was better able to recognise inverted faces than he could upright faces suggesting that there is indeed an upright face recognition region in the brain.
One explanation which may account for why the normal test subjects were better able to recognise upright faces comes from Diamond and Carey (1986) who proposed that configural information, such as the distance between features of the face, for example the eyes, is used to distinguish between faces.
Inverted faces may make it harder to process such information thus reducing ones ability to recognise an inverted face they have seen before, as has been famously demonstrated by the “Thatcher illusion” (Thompson, 1980).
Aims
The aim of this experiment was to determine whether or not there is a special face processing system by comparing the processing of upright and inverted faces, with the processing of upright and inverted landscapes.
Hypothesis
The experimental hypothesis is that stimulus inversion will impair the recognition of face stimuli more than landscape stimuli.
Method
Design
This experiment was of a between subjects design as participants were allocated to different groups (i.e. face or land). The independent variable which was manipulated was the presentation of face and landscape images upright or inverted to participants in the face or land group. The dependent variable which was measured was the correct scores given by participants for the target images.
Participants
In this experiment 12 males and 12 females aged 18-40 were randomly allocated to either a “face” or “land” group, with each group containing 12 participants. All participants were Level 2 psychology students at the University of Aberdeen, Scotland.
Materials
For the learning phase of the experiment, 12 upright and inverted male face images were used for the face group, and 12 upright and inverted landscape images were used for the land group.
For the testing phase of the experiment, 24 images of upright and inverted male faces were used for the face group, and 24 images of upright and inverted landscapes were used for the landscape group. All images were displayed via an overhead projector onto a projector screen using Microsoft PowerPoint.
Of the 24 images used for the face and land groups, 12 were target images and 12 were distracter images.
Response sheets listing each image from 1-24 were used to record participants memory of target upright and target inverted images.
Procedure
Participants were randomly allocated into either a face group, who were shown images of faces, or a land group, who were shown images of landscapes.
The initial phase of the experiment began with a learning phase which involved presenting 12 upright face images to the face group, and 12 upright landscape images to the land group for 1 second per image.
The testing phase of the experiment involved presenting 12 upright face images to the face group, and 12 upright landscape images to the land group for 6 seconds per image.
After a 30 second pause, participants were then shown 24 upright face images for the face group, and 24 upright landscape images for the land group. Participants were instructed to write down on their response sheet whether they had previously seen the image, a yes response, or whether they had not seen the image, a no response.
During this phase of the experiment, 12 of these 24 images consisted of target images which the participants had previously been exposed to. The other 12 images were distracter images which the participants had not previously been exposed to.
After completion of the upright image phase of the experiment, subjects were then shown inverted images using the same procedure that was used for the upright images including a learning phase for inverted images.
Once the experiment had been completed, the experimenter collected the response sheets from all participants and then gave all participants a main analysis data sheet with upright and inverted scores for the face and land group alongside difference scores for each participant and a mean difference score for faces and for landscapes (see appendix).
Results
Group | Mean Upright Score | Mean Inverted Score |
| Face | 10.9 | 8.50 |
| Land | 10.3 | 10.2 |
Table 1
– Mean upright and inverted scores for face and land main data groups.
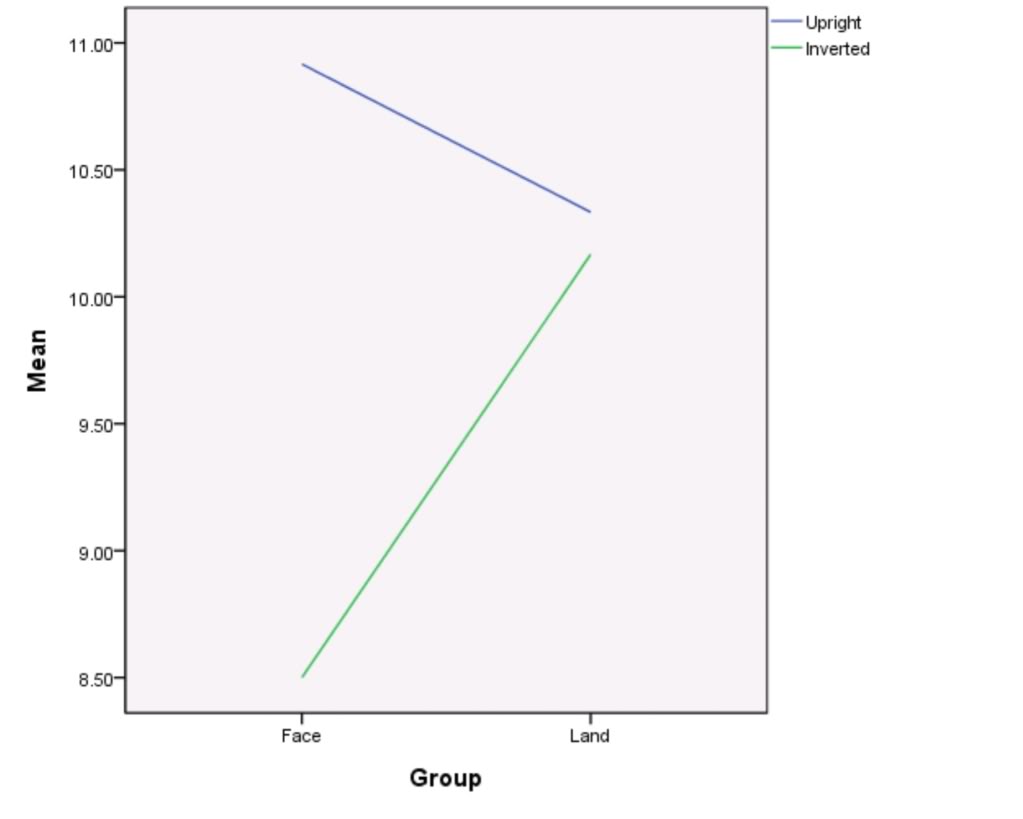
Figure 1
– Line graph to show mean upright and inverted scores for face and land main data groups.
As shown by table 1 and figure 1, the mean upright score from the main face group data was 10.9 and the mean upright score from the main land group data was 10.3. These results differ by a score of 0.60.
The mean inverted score for the main face group data was 8.50 and the mean inverted score for the main land group data was 10.2. These results differ by a score of 1.70.
The difference between the mean upright score and the mean inverted score for the main face group data was 2.40. In contrast, the difference between the mean upright score and mean inverted score for the main land group data was 0.10.
Table 2
– Mean upright and inverted scores for face and land class data groups.
Group | Mean Upright Score | Mean Inverted Score |
| Face | 10.0 | 8.62 |
| Land | 8.48 | 9.64 |
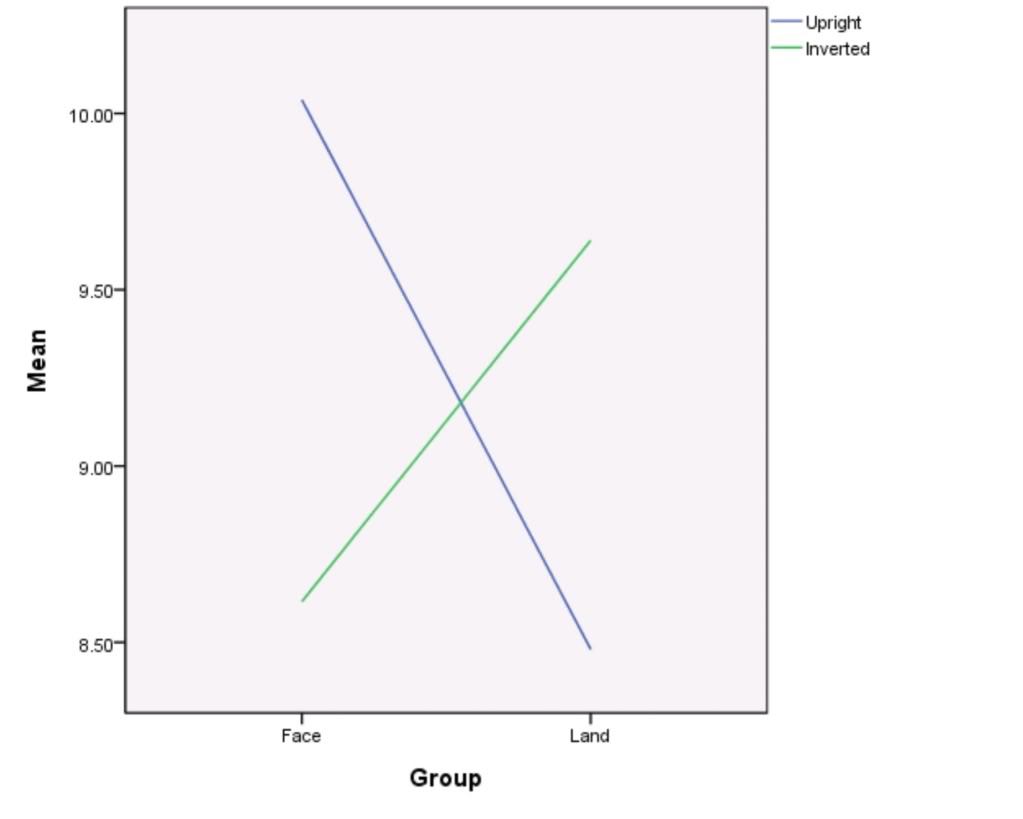
Figure 2
– Line graph to show mean upright and inverted scores for face and land class data groups
As shown by table 2 and figure 2, the mean upright score from the class face group data was 10.0 and the mean upright score from the class land group data was 8.48. These results differ by a score of 1.52.
The mean upright score from the class face group data differs from the mean upright score from the main face group data by a score of 0.90. The mean upright score from the class land group data differs from the mean upright score from the main land group data by a score of 1.82.
The mean inverted score for the class face group data was 8.62 and the mean inverted score for the class land group data was 9.64. These results differ by a score of 1.02.
The mean inverted score from the class face group data differs from the mean inverted score from the main face group data by a score of 0.12. The mean inverted score from the class land group data differs from the mean inverted score from the main land group data by a score of 0.56.
The difference between the mean upright score and the mean inverted score for the class face group data was 1.38. In contrast, the difference between the mean upright score and mean inverted score for the class land group data was 1.16.
An independent-samples t-test was carried out on the main face and land group difference scores in order to examine the experimental hypothesis.
The results from the independent-samples t-test showed that the effect of inversion for faces (M = 2.42, SD = 1.78) was greater than that for landscapes (M = .17, SD = 1.85): t (22) = 3.03, p < .006. As the p value is less than the critical value of .05, it can be said that inversion has a statistically significant effect on the recall of target images and that these results were unlikely to have been obtained by chance.
In addition, because the mean difference score for faces (2.42) was greater than the mean difference score for landscapes (.17), it can be said that inversion had a greater effect on the recall of target face images as opposed to target landscape images.
This data therefore supports the experimental hypothesis that stimulus inversion will impair the recognition of face stimuli more than landscape stimuli.
Discussion
The aim of this experiment was to determine whether or not stimulus inversion impairs the recognition of face stimuli more than landscape stimuli.
As shown by the mean difference scores, the effect of inversion was indeed greater for face images (mean difference score = 2.42) than it was for landscape images (mean difference score = .17). The recorded p value of .006 was less than .05 indicating that these results were statistically significant. The data obtained from this experiment therefore provides statistically significant support for the experimental hypothesis allowing for it to be accepted.
The results obtained from this experiment are similar to the results obtained from prosopagnosics such as patient W.J (McNeil et al, 1993) and patient B.M (Sergent et al, 1989) in the sense that they suggest that there may be a region of the brain which is involved in facial recognition. This was shown by inversion having a greater adverse effect on the recognition of facial images than it did for the recognition of landscape images.
These findings, in addition to the findings from prosopagnosics P.S (Rossion et al, 2003) and D.F (Steeves et al, 2006), both of whom who had a damaged OFA but were still able to recognise objects, suggests that this face recognition region may in fact be solely devoted to the recognition of faces and is not needed for the recognition of other stimuli.
However, it is important to note that some prosopagnosics also suffer from impaired object recognition (Farah, Levinson, and Klein, 1995), which would mean that it also processes objects to some degree.
Furthermore, the ability of prosopagnosic patient L.H to recognise inverted facial images better than upright facial images (Farah et al, 1995), also suggests that this face recognition region is specifically designed to process upright faces rather than just faces in general.
From an evolutionary perspective this makes sense, as from two months of age babies begin to preferentially process faces at the expense of other stimuli (Johnson, Dziurawiec, Ellis and Morton, 1991) and the faces they see are, for the most part, always in an upright orientation.
Whether or not this preference for upright stimuli is an inborn characteristic remains unknown. It may be the case that the brain merely adapts itself to the predominant orientation of stimuli in its environment and that if babies only saw inverted stimuli from birth they would, as adults, perform better at recognising inverted faces and worse at recognising upright faces. In effect, producing results opposite to those found in this experiment.
Of course, for ethical reasons this is something that we are unlikely to ever find out. However, the idea of an orientational adaptability of the brain is not beyond the realms of possibility.
For example, research carried out by Blakemore and Cooper (1970) on kittens who were raised from birth in an environment with only vertical stripes, showed that when placed into an unrestricted visual environment the kittens could not see horizontal lines (Grobstein and Chow, 1975) as their visual cortex had adapted itself to the predominant orientation (vertical) of stimuli that was present in their environment since birth.
In addition to the orientation of an image, it is also important to consider the configurational information that lies within that image. It has been shown for example, that the recognition of dog faces is as sensitive to inversion as human faces, providing that one is an expert at recognising the breed of a dog (Diamond et al, 1986).
Since human faces and the faces of a particular breed of dog share similar configurational information, such as the distance between the eyes, nose and mouth between different humans, or the distance between the eyes, nose and mouth between different dogs within the same breed, this may partly explain why the recognition of faces is so adversely affected by inversion as once this information becomes distorted, such as through inversion, it also distorts our ability to recognise a face as that information can no longer be used, or is more difficult to use, for recognition.
In contrast, the recognition of landscape images, which do not share such similar configural information between different landscapes, is much less sensitive to the adverse effects of inversion.
This may be because configural information is not used for the recognition of such images, or is used to a lesser extent, and so inversion distorts the information in landscape images less than it does for facial images and so has less of an effect on recognition.
Thus from these findings we can infer that yes, there may be a specific face recognition region of the brain, and that it appears to preferentially process upright face images using configural information for the recognition of a particular face.
One interesting observation to note from this experiment is the scores obtained from the land group class data. In contrast to the land group main data which achieved a mean upright score of 10.33 and a mean inverted score of 10.17, the class data achieved a mean upright score of 8.48 and a mean inverted score of 9.64, showing that participants performed better in the inverted scenario whereas participants in the main group performed better in the upright scenario.
One possible explanation for why this may have occurred is that participants in the class land group may have benefited somehow from seeing images upright during the first testing phase.
They may for example, gained a better understanding of what they were required to do in the experiment after seeing the upright images, thus priming them for the next testing phase with inverted images.
To help eliminate or reduce this possible confounding variable, future experiments should utilise counterbalancing to reduce such practice effects. This could be done by showing the face group upright images first and inverted images second, and the land group inverted images first and upright images second.
It is also difficult to make an accurate comparison between the main group data and the class group data as different sample sizes were used. With the main group data 12 participants took part in the face group and 12 participants took part in the land group. An equal allocation of participants between groups.
However, with the class data, 26 participants took part in the face group and 25 participants took place in the land group. An unequal allocation of participants between groups.
As there were fewer participants in the class land group, this may have influenced the mean scores obtained. Future experiments would therefore benefit from having an equal allocation of participants between the two groups.
Another consideration worth taking into account is the type of participants who made up each group as they were made up of a mixture of male and female participants.
Some studies indicate that females perform better at face recognition tasks than males (Bigun and Smeraldi, 2001), thus if there was an unequal distribution of males and females in the class face group and class land group, this may have influenced the scores obtained by each group. Future experiments may therefore benefit from having female only face and land groups, which could then be compared to male only face and land groups.
The effect of the sex of the face images participants are exposed to could also be investigated in future experiments. In this experiment, participants were shown only male faces.
However, this may have biased the results somewhat as some studies have shown that women are better at recognising female faces, and men are better at recognising male faces (Lewin and Herlitza, 2002).
Further experiments could therefore test the face recognition abilities of men for male and female faces, and the face recognition abilities of women for female and male faces.
Furthermore, not all images of faces shown in this experiment had the same hair length or hair style which may have affected face recognition (Wright and Sladden, 2003).
Future experiments could be carried out groups who are shown only one type of hairstyle/hair length, and then compared to other groups who were shown faces with another type of hairstyle/hair length.
Finally, the race of the face images participants were shown and the race of the participants themselves may have acted as another confounding variable, as some studies have shown that face recognition tends to be better for own race faces (Tanaka, Kiefer and Bukach, 2004). Future experiments could therefore investigate the effects on facial recognition of showing Caucasians Caucasian faces and Asians Asian faces for example.
Conclusion
The results of this experiment showed that image inversion disrupted the processing of face stimuli more than the processing of landscape stimuli.
As a result, the proposed experimental hypothesis, that stimulus inversion will impair the recognition of face stimuli more than landscape stimuli, can be accepted. On the basis of these findings, there does appear to be a specialised face processing region in the brain.
References
Barrera, M., & Maurer, D. (1981). The perception of facial expressions by the three-month-old. Child Development 52, 203-206.
Bigun, J., & Smeraldi, F. (2001). Evidence on skill differences of women and men concerning face recognition. Audio and Video Based Biometric Person Authentication Lecture Notes in Computer Science 2091, 44-50.
Blakemore, C., & Cooper, G. (1970). Development of the brain depends on the visual environment. Nature 228, 477-478.
Diamond, R., & Carey, S. (1986). Why faces are and are not special: An effect of expertise. Journal of Experimental Psychology 115, 107-117.
Duchaine, B., & Nakayama, K. (2005). Dissociations of face and object recognition in developmental prosopagnosia. Journal of Cognitive Neuroscience 17, 1-13.
Farah, M.J., Levinson, K.L., & Klein, K.L. (1995). Face perception and within-category discrimination in prosopagnosia. Neuropsychologia 33, 661-674.
Farah, M.J., Wilson, K.D., Drain, H.M., & Tanaka, J.R. (1995). The inverted face inversion effect in prosopagnosia: Evidence for mandatory face-specific perceptual mechanisms. Vision Research 35, 2089-2093.
Grobstein, P., & Chow, K.L. (1975). Receptive field development and individual experience. Science 190, 352-358.
Johnson, M.H., Dziurawiec, S., Ellis, H., & Morton, J. (1991). Newborns’ preferential tracking of face-like stimuli and its subsequent decline. Cognition 40, 1-19.
Kurucz, J., & Feldmar, G. (1979). Prosopo-affective agnosia as a symptom of cerebral organic disease. Journal of the American Geriatrics Society 27, 225-230.
Langlois, J.H., Ritter, J.M., Roggman, L.A., & Vaughn, L.S. (1991). Facial diversity and infant preferences for attractive faces. Developmental Psychology 27, 79-84.
Lewin, C., & Herlitza, A. (2002). Sex differences in face recognition: Women’s faces make the difference. Brain and Cognition 50, 121-128.
Maurer, D., & Barrera, M. (1981). Infants’ perception of natural and distorted arrangements of a schematic face. Child Development 52, 196-202.
McNeil, M., & Warrington, E.K. (1993). Prosopagnosia: A face-specific disorder. Quarterly Journal of Experimental Psychology 46, 1-10.
Rossion, B., Caldara, R., Seghier, M., Schuller, A.M., Lazeyras, F., & Mayer, E. (2003). A network of occipito-temporal face-sensitive areas besides the right middle fusiform gyrus is necessary for normal face processing. Brain 126, 1-15.
Samuels, C.A., & Ewy, R. (1985). Aesthetic perception of faces during infancy. British Journal of Developmental Psychology 3, 221-228.
Sergent, J., & Villemure, J.G. (1989). Prosopagnosia in a right hemispherectomized patient. Brain 112, 975-995.
Steeves, J.K., Culham, J.C., Duchaine, B.C., Pratesi, C.C., Valyear, K.F., Schindler, I., Humphrey, G.K., Milner, A.D., & Goodale, M.A. (2006). The fusiform face area is not sufficient for face recognition: evidence from a patient with dense prosopagnosia and no occipital face area. Neuropsychologia 44, 594-609.
Tanaka, J.W., Kiefer, M., & Bukach, C.M. (2004). A holistic account of the own-race effect in face recognition: evidence from a cross-cultural study. Cognition 93, 1-9.
Thompson, P. (1980). Margaret Thatcher: a new illusion. Perception 9, 483-484.
Wright, D.B., & Sladden, B. (2003). An own gender bias and the importance of hair in face recognition. Acta Psychologica 114, 121-128.